 EDITORIAL
EDITORIAL
Parvaiz Ahmad1,2, Javaid Akhter Bhat3, Luisa María Sandalio4, Muhammad Ashraf5
1Botany and Microbiology Department, King Saud University, Riyadh, Saudi Arabia
2Department of Botany, Degree College Pulwama, Jammu and Kashmir, India; parvaizbot@yahoo.com
3State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, China javid.akhter69@gmail.com
4Bioquímica, Biología Celular y Molecular de Plantas, Estación Experimental del Zaidín, CSIC Albareda 1, 18008 Granada, Spain luisamaria.sandalio@eez.csic.es
5Depatment of Botany, University of Agriculture Faisalabad, Pakistan; ashrafbot@yahoo.com
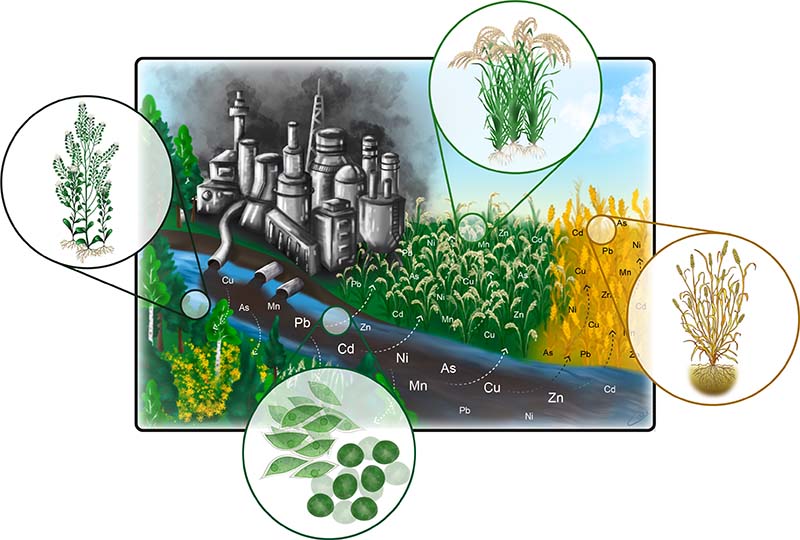
The levels of heavy metals (HMs) have increased tremendously in the environment over the past decades, and they have caused considerable problems for the world community (Ashraf et al., 2019). Metal elements with high molecular mass and density are included in the HMs, such as copper (Cu), zinc (Zn), nickel (Ni), lead (Pb), cadmium (Cd), chromium Cr) and mercury (Hg) (Bhat et al. 2020). The HMs presence in soil occurs naturally or through human activities (as reviewed in detail by Briffa, 2020; Peco et al., 2021). These HMs remain in intact form in the ecosystem for a long time as they are not degradable by physico-biological processes; they are thus a great threat to the ecosystem (Suman et al., 2018). Based on their function in plants, HMs are categorized into essential and non-essential. For example, Zn, Fe, Cu, Ni and Mn are essential for regulating the optimal growth and development in plants (Cempel and Nikel, 2006). However, at higher concentrations, they cause adverse effects to plants. On the other hand, the non-essential HMs such as Cd, Hg, and As cause highly toxic effects on plants even at low levels (Fasani et al., 2018). These toxic HMs can get into the human via the food chain from the crop plants, leading to hazardous effects on human health (Rahman et al., 2019).
There are direct or indirect toxic effects of HMs on plants. The direct negative effects of the HMs on plants include enzyme inhibition and oxidative stress-induced injury (Bhat et al. 2020). The HM-mediated indirect toxic effects on plants occur due to HM-mediated replacement of essential nutrients from the cation exchange sites. A decrease in plant growth under HM stress is the consequence of mineral homeostasis disturbances and the reduction of photosynthesis and antioxidant defense activities (Rabelo et al. 2021; Alamri et al. 2021). Besides, HM toxicity influences the beneficial microorganisms of soil, indirectly affecting plant growth (Morkunas et al. 2018). Although different approaches have been used previously to reclaim HM polluted soils, these measures are based on chemical, physical and mechanical techniques (as reviewed by DalCorso et al., 2019; Peco et al., 2021). These methods possess many demerits limiting their use on a large scale. Such demerits include expensiveness, reduced efficiency for removal of metals at low concentration, disturbance of the normal biological and physio-chemical processes of soils, destruction of the soil bio-network, and insertion of secondary pollutions (DalCorso et al., 2019; Peco et al., 2021). Therefore, the scientific community intends to adopt approaches that are eco-friendly, efficient, and cost-effective. In this regard, an approach based on plant system, called phytoremediation, has emerged. In this approach, plants are exploited to remove HMs from the contaminated soil or reduce their availability in soil for plant uptake (Jacob et al., 2018; Alam et al. 2021; Nigam et al. 2021; Peco et al., 2021). The use of phytoremediation for the reclamation of the soils polluted with HMs has many merits, such as being cost-efficient, environmentally safe, easy to manage, large scale application, and improved soil fertility (Wuana and Okieimen, 2011; Jacob et al., 2018; Majeed et al. 2021). Several investigations have been carried out during the last few decades to explore the molecular mechanisms involved in HM tolerance and establish protocols for improving the efficiency of phytoremediation (Kaya, 2021; Nigam et al. 2021; Souri et al. 2021; Kabir et al. 2021; Jogawat et al. 2021; Zahra et al. 2021; Rana et al. 2021). The phytoremediation mechanisms include phytoextraction, pytodegradation/phytotransformation, phytovolatilization, phytostabilization and rhizofiltration.
Some plants possess the capability to accumulate high levels of the HMs in their shoot tissues, i.e., > 0.1% of dry weight and therefore can be useful for phytoextraction (Susarla et al. 2002). This process can be carried out by plants via two ways, i.e., continuous phytoextraction and induced phytoextraction. Plants possessing a natural ability to accumulate a high quantity of HMs (hyperaccumulators) can effectively carry out continuous phytoextraction (Sytar et al. 2021). However, in the case of induced phytoextraction, different types of chemicals, such as chelates or plant signalling compounds such as nitric oxide (Souri et al. 2021), 2-oxoglutarate (Alamri et al. 2021), 24-epibrassinolide (da Silva Cunha et al. 2021) and hydrogen sulphide (Kaya et al. 2021), can enhance the plants’ ability to accumulate more HMs (McGrath et al. 2002). Plant species such as Sesbania exaltata and Sesuvium portulacastrumi, possessing deep root system and higher growth rate (an important trait for phytoextraction), as well as some microalgae (e.g., Scenedesmus acutus and Chlorella pyrenoidosa) have been documented to have great potential for HM phytoextraction (see details at Nedjimi, 2021; Shivagangaiah et al. 2021; Zhang et al. 2021). The phytodegradation/phytotransformation approach involves the use of metabolic processes to degrade HMs/pollutants by plants, either within or outside the plants (Schnoor et al. 1995). For example, Rugh et al. (1998) found that genetically modified Liriodendron tulipifera (yellow poplar) transforms Hg2+ to a less toxic form of mercury, i.e., Hg. This method allows the plants to convert toxic pollutants into non-toxic ones (Rai et al. 2021). Phytovolatilization, an important approach of HMs detoxification, transforms the absorbed toxic pollutants into non-toxic volatile forms. This method works via the initial absorption of metals by plant roots, and their subsequent transport to the shoot, and ultimately their volatilization into the atmosphere via the leaf stomata (Tollsten and Muller, 1996). For example, Bañuelos et al. (1996) revealed the role of the Indian mustard for Se phytovolatilization from the polluted soil. Similarly, other plants have also been used for phytovolatilization, such as Brassica juncea (Moreno-Jiménez et al. 2009), Nicotiana tabacum (Ashraf et al. 2010) and Arundo donax (Guarino et al. 2020). Another approach, ‘rhizofiltration’, operates via the precipitation from the water/liquid waste on the root surface or by root absorption of soluble pollutants (Dushenkov et al. 1995). In this approach, a root system with maximum surface area, such as fibrous root system, has been found to be more relevant (Salt et al. 1998). Besides, the plant’s ability to absorb a large quantity of water from the soil is also significant in this approach (Susarla et al. 2002). This strategy of metal detoxification has been recorded in plant species such as Phaseolus vulgaris (Yang et al. 2015), Arundo donax (Oustriere et al. 2017), Salvinia molesta and Pistia stratiotes (Kodituwakku and Yatawara 2020). Phytostabilization is another strategy used by plants for the removal of HMs from polluted soils; in this approach, plant roots store the HM pollutants or HMs are precipitated as root exudates (Peer et al. 2005). This approach reduces the pollutant movement, thereby preventing their transfer into groundwater. For example, Nedjimi and Daoud (2009) revealed that Atriplex halimus plants possess a high ability for the phytostabilization of Cd thanks to a suitable root architecture. The phytostabilization ability has also been reported in plant species such as Erica australis (Monaci et al. 2019), Helichrysum microphyllum (Bacchetta et al. 2018), Pelargonium hortorum (Manzoor et al. 2020), Quercus robur (Susarla et al. 2002) and Salix alba (Mataruga et al. 2020).
Besides, some advanced approaches for refining phytoremediation are also available such as microbial‐assisted phytoremediation (PGPR), AMF inoculation‐assisted phytoremediation, earthworm‐assisted phytoremediation, phytohormone‐assisted phytoremediation, nanoparticles‐assisted phytoremediation and transgenic approaches. Plant growth-promoting bacteria (PGPB) present in the rhizosphere zone possess the ability for bioremediation. These bacteria allow the plants to improved their growth and mineral nutrition. Such bacteria either transform the toxic HMs into non-toxic components or they degrade the toxic pollutants (Ullah et al. 2015; Ummara et al. 2021). Some studies have documented multiple numbers of PGPB that improve the ability of plants to phytoremediate, especially by allowing higher HM uptake of plant roots. By secreting substances in the soil, such as organic acids and siderophores (chelators), bacteria improved HM detoxification; these substances increase HM bioavailability via reducing the pH of the soil (Chen et al. 2017). Many bacteria belonging to different strains enhance plant tolerance to HMs (as reviewed by Nedjimi, 2021). Furthermore, fungi symbiosis with plant roots, in the form of arbuscular mycorrhizal fungi (AMF), enhances the phosphorus availability for plant uptake (Zhang et al. 2020). Decontamination of HMs by AMF occurs via two pathways, (1) AMF-producing chelators immobilizing HMs, and their adsorption to cell wall of fungi, and (2) they enhance the growth of plants by increasing the phytoextraction of HMs via modulating the pH of soil or the chemical composition of the root exudates (Cabral et al. 2015). Earthworms are living organisms belonging to macroinvertebrates, and they are often called the engineers of the ecosystem. These earthworms decompose soil organic matter; besides, they allow the cycling of nutrients as well as enhance the soil quality (Sharma et al. 2020). These organisms secrete fulvic and humic acids (Lemtiri et al. 2016; Wang et al. 2020), which change the pH of soil that in turn increases the bio-availability of HMs and nutrients. Plant growth regulators (PGR)-mediated phytoremediation allows enhanced accumulation of HMs in plant tissues. In this regard, four major phytohormones have been reported to play a role in the phytoremediation of HMs: auxins (IAA), cytokinins, gibberellins (GAs) and abscisic acid (ABA). Some studies have documented that these hormones enhance HM accumulation as well as HM tolerance and growth in plants. Phytohormone application to plants especially at earlier growth stage allows the plants to escape toxicity under HM stress conditions. It has been shown for auxins in Arabidopsis thaliana (Zhu et al. 2019), gibberellic acid 3 (GA3) in Solanum nigrum (Ji et al. 2015) and ABA in Vitis vinifera (Song et al. 2019). Recently, nanoparticles (NPs) have emerged as a novel method of improving the plants’ efficiency for HM removal (Zhu et al. 2019). The NPs increase the ability of plants for phytoremediation in different ways: (1) NPs interact with HMs via redox reactions/adsorption, (2), NPs stimulate plant growth, and (3) assist the phytoremediation of HMs (Song et al. 2019). Interaction at the chemical level revealed that NPs assist the plants for HM stabilization via electrostatic adsorption. Several works have demonstrated the beneficial impact of nanoparticles in the form of increased phytoremediation of HMs such as Ag nanoparticles (AgNPs) (Khan and Bano 2016), nano-TiO2 particles (Singh and Lee, 2016), nanoscale zero-valent iron (nZVI) (Gong et al. 2017; Huang et al. 2018), and magnesium oxide (MgO) nanoparticles (Hussain et al. 2019).
Transgenic plants possess foreign gene/genetic elements that improve plant efficiency to uptake and translocate HMs (Rai et al. 2020). This approach is now considered an important biotechnological innovation for phytoremediation improvement. The overexpression of genes, from plants and microbes, was utilized to decrease the HM stress as well as enhance the plants’ ability to remove HMs from polluted areas (Liu et al. 2020). The use of transgenic plants in HM detoxification relies on the objective to enhance the plant capacity for HM uptake, accumulation, as well as their degradation and tolerance (Gomes et al. 2016). Besides, this technique enhances the agronomic properties of plants for achieving enhanced growth, biomass, yield and well-organized root system, with a rapid growth in different soil types (Rai et al. 2020). For instance, the mercuric ion reductase (merA) and organomercurial lyase (merB) are the two bacterial genes overexpressed in the plant system to efficiently detoxify Hg (Bizily et al. 2003). The merB gene catalyses the conversion of the organic form of Hg to inorganic ionic form of Hg2+ that is far less toxic than the organic one (Bizily et al. 2003). Similarly, Chaturvedi et al. (2014) demonstrated that SbMT-2 gene, discovered from Salicornia brachiate, can mediate detoxification of HMs (Zn, Cd and Cu), thereby leading to HM tolerance, especially by modulating the ROS elimination in transgenic Nicotiana tabacum. Moreover, this approach has been used for HM detoxification by many other researchers in different plants (Peng et al. 2017; Meena et al. 2018; Kumar et al. 2019; Upadhyaya et al. 2021; Ahammed et al. 2021; Sanz-Fernandez et al. 2021; Khoudi, 2021; Gomez Mansur et al. 2021).
The above discussion shows the important role of phytoremediation in the decontamination of HM polluted soils. However, more research efforts are required. Besides, to enhance the efficiency of the plant-based elimination of HMs from the polluted regions, it is a prerequisite to elucidate the underlying mechanisms at the physiological, biochemical and molecular levels, especially by using the recent advanced ‘Omics’ approaches. Moreover, the interaction among various phytoremediation approaches are required to expand the prospects of polluted soil remediation. Lastly, the successful use of phytoremediation depends squarely on the cooperation and joint efforts of the farming and local communities, scientists, different sectors of industry and authorities of environmental control by conducting educational programs, thus allowing long-term sustainability of this friendly green technology.
The present special issue covers recent advances made in the sequestration of HMs from the contaminated and polluted areas as well as confirms the role of plants and microbes in the decontamination of HM-polluted regions, and the underlying mechanisms of this HM removal. The transgenic approaches, nanotechnology and other plant-based approaches used to improve plant efficiency for the clean-up of contaminated soils, as well as to enhance HM tolerance of plants, are also present in this Special Issue. We hope this special issue provides an excellent set of papers for a greater understanding of HM sequestration from the contaminated environments as well as HM tolerance in plants with novel ideas for future research and development on metal stress biology.